Fertiliser
Organic Veg Fertiliser 4-4-4 | Leafy Growth
Balanced 4-4-4 for strong leafy growth.
from £10.99
Free UK Delivery on Orders Over £40









Dr Forest
Couldn't load pickup availability
Dr Forest customers rate us 4.89/5 · 3,250+ reviews across Shopify, Amazon, Google and eBay
Bloom 2-8-4 is a high-phosphorus organic bloom fertiliser — a natural bloom booster built for the flowering and fruiting stage. The ratio is phosphorus-dominant — 8% P₂O₅ drives flower initiation, bud development, and fruit set, while reduced nitrogen at 2% prevents the excess leaf growth that thins out flowers. Potassium at 4% supports sugar transport into developing buds and fruit, sulphur runs at over 5% to feed the terpene and aroma compounds that define a quality harvest, and calcium sits high near 8% for cell wall construction during fruit set. Twenty organic and mineral inputs deliver this in one handcrafted granular blend.
This is a living-soil flower fertiliser. Nutrients release through microbial activity, not chemical solubility — the same biology that produces the secondary metabolites responsible for flavour, aroma and resin in the finished crop. Bloom 2-8-4 is our bestselling bloom feed and the natural partner to Veg 4-4-4 for growers who split their feeding between vegetative and flowering stages. Handcrafted in small batches in Stockport.
We make three bloom fertilisers, each built from the same organic inputs in different proportions. Bloom 2-8-4 is our bestseller — high phosphorus, moderate potassium, ideal as a standalone bloom feed or for early-to-mid flower. Bloom 2-4-8 shifts the balance toward potassium for growers who favour higher K in the mid-to-late flowering phase for sugar transport and ripening. Bloom 2-8-10 delivers high P and high K together — for heavy-feeding varieties or growers who want maximum phosphorus and potassium throughout. Many growers use a single bloom all the way through; others run 2-8-4 into early flower then switch to 2-4-8 for the finishing weeks. Both work — pick the ratio that matches how you grow.
If you are running separate veg and bloom fertilisers rather than one product through the whole grow, you are already growing for maximum results. We built Bloom 2-8-4 for that approach — and we loaded it with the expensive ingredients that drive results, which is why it costs more than our Premium range (Tomato, Rose & Flower, Fruit & Veg, Strawberry). Compared to those blends, Bloom 2-8-4 contains 2.5× the humic and fulvic acid and double the Scottish seaweed. Veg and Bloom also include diastatic malted barley and the Amino Acid Powder we sell as a standalone — neither of which is in any of our other fertilisers. The Premium range is excellent for one-product growers. Bloom 2-8-4 is for growers who want the very best at each stage.
Every batch is blended by hand in our Stockport unit and made with organic ingredients throughout, with British sources used wherever we can find them — including Scottish seaweed. No slaughterhouse waste, no synthetic chemistry.
Bloom 2-8-4 carries the same premium ingredient set as our Veg 4-4-4, blended in different proportions to deliver a phosphorus-dominant bloom ratio. Multiple sources stack at each nutrient — three for phosphorus, three for potassium, three for calcium, four for sulphur — so the plant draws steadily from a deep nutrient bench through the entire flowering cycle. Reduced nitrogen prevents the vegetative stretch that thins flowers.
British-produced plant protein concentrate at 12% N — included at a deliberately reduced rate for the bloom stage. Provides background nitrogen to support leaf function and the early flower stretch without driving the soft, airy growth that excess N produces during flower. Releases gradually through microbial mineralisation.
The primary phosphorus source — a calcined plant meal at roughly 15% P₂O₅ and 9% CaO. The phosphorus is citric acid soluble, which means plant roots can unlock it on demand through their own organic acid exudates. During flower bud differentiation and fruit set the plant secretes citrate and malate into the rhizosphere, dissolving the meal and releasing P directly into the root zone where it is needed.
A high-grade plant-derived free amino acid concentrate at 12% N — the same Amino Acid Powder we sell as a standalone product on the website. Free amino acids are absorbed by plant roots in pre-formed nitrogen units, bypassing the energy-intensive nitrate-to-amino-acid conversion that takes place inside the plant. Particularly useful during flower, when carbon budgets are tight and every unit of metabolic energy spent on nitrogen assimilation is energy not spent on flower production.
Cold-milled lucerne containing triacontanol — a natural long-chain alcohol that increases photosynthetic efficiency. Photosynthesis is the limiting resource during flower; every additional percentage point translates into more sugars available for sinks. Alfalfa breaks down quickly in moist soil, releasing background N and K plus growth-promoting compounds across the first three weeks of bloom.
The primary potassium source — K₂SO₄ at 50% K₂O and 45% SO₃. Chloride-free, which matters: chloride salts suppress flavour compound accumulation and damage soil biology over repeated cycles. Potassium regulates stomatal opening and the long-distance transport of sucrose from leaves to flowers and fruit. Sulphur is a precursor for the sulphur-containing volatile compounds that anchor aroma.
Ultra-finely milled apatite mineral at 31% P and 49% Ca. The second phosphorus source in the blend, providing a slow-release reserve that continues feeding flowers long after faster sources are depleted. Micronisation increases the surface area exposed to soil acids and root exudates, lifting availability well above coarse rock phosphate. The high calcium content underpins the elevated total Ca in the finished blend.
Calcium sulphate dihydrate (CaSO₄·2H₂O) at 23% Ca and 17% S. Soluble calcium without the pH lift of limestone — important for established living soil that already sits in the right pH band. Calcium loads cell walls during the rapid expansion of flower and fruit tissue; the sulphate fraction supports cysteine and methionine synthesis, both essential for terpene precursor production.
Magnesium sulphate monohydrate at roughly 25% Mg and 50% S. Soluble magnesium pairs neatly with the calcium load — magnesium sits at the centre of every chlorophyll molecule, and chlorophyll is the engine that powers flower production. The high sulphate contribution keeps the blend's S budget supplied for amino acid and terpene biosynthesis.
A nutrient cycling activator unique to our Veg and Bloom blends. Germinated barley containing active amylase and protease enzymes that break down starches and proteins in the soil into plant-available sugars and amino acids. Accelerates the microbial cycling that releases secondary metabolites responsible for flavour and aroma. Not in any of our other fertilisers.
Pyrolysed carbon with internal surface area approaching 300 m² per gram, pre-charged with microbial communities through fermentation. Bio-char acts as permanent microbial habitat — critical during flower, when steady microbial activity drives the secondary metabolite production that determines crop quality. Lehmann et al. (2011) showed K retention increases of 18–35% under leaching conditions.
Humic acid lifts cation exchange capacity, builds aggregate structure and stimulates root growth. Fulvic acid is a low-molecular-weight chelator that binds micronutrients into plant-available complexes and shuttles them across cell membranes — vital during flower when micronutrient demand for enzyme cofactors peaks. Bloom 2-8-4 carries this at 2.5%, around 2.5× the level in our Premium range.
Cold-water Ascophyllum hand-harvested from the Scottish Atlantic coast. Contains cytokinins, auxins, gibberellins, alginic acid, laminarin and over 60 trace elements. During flower the cytokinin activity delays leaf senescence, keeping the canopy photosynthetically active longer to power continued flower and fruit development. The whole-plant powder runs at 4% — double the level in our Premium range — and pairs with the concentrated seaweed extract further down the list for a fast-acting plus slow-release biostimulant pairing.
Medium-speed nitrogen source from cold-pressed oilseed rape. Contributes background N, a small phosphorus fraction and modest sulphur from the residual sulphur compounds in the seed. The slow protein breakdown extends the nitrogen tail across the early-to-mid flower window without spiking levels at any single point.
Insect castings rich in nitrogen, phosphorus, potassium and chitin. Chitin triggers the plant's systemic defence response through a well-documented elicitor pathway — particularly valuable during flower, when pest pressure tends to rise and the crop is at its most valuable. Frass also contributes a biological phosphorus fraction that microbes mineralise steadily through the bloom cycle.
High cation exchange capacity (CEC) clay minerals that hold positively charged nutrient ions and release them gradually to plant roots. During flower, nutrient uptake fluctuates rapidly — clay acts as a buffer, holding the line through demand spikes and protecting against the deficiency swings that come from passing showers or heavy waterings.
A blend of beneficial soil bacteria and yeast cultures introduced as a dormant powder. Once activated by water, the inoculant boosts the breakdown of organic matter, supports the production of plant-available nutrients and contributes to a more diverse rhizosphere community. New to the formulation — particularly useful when amending used substrate between cycles.
A proprietary blend of dried herbs and botanical material contributing trace plant compounds, plant-derived growth factors and additional carbon forms for soil biology to work on. Broadens the range of organic substrates available to the microbial community, supporting the metabolic diversity that drives quality compound production through the bloom cycle.
Volcanic basalt ground to a fine powder, carrying over 60 trace elements — iron, manganese, zinc, copper, boron, molybdenum, cobalt, selenium. Trace elements are critical during flower: boron drives pollen tube growth, zinc regulates auxin synthesis, manganese supports photosynthesis, iron underpins chlorophyll production. Released slowly over several years, building the trace mineral reserve in the substrate.
Plant-available silicon — the structural nutrient that strengthens cell walls and increases stem rigidity. During flower, silica-fed plants produce thicker stems capable of supporting heavy bud and fruit loads without staking. Silicon also improves resistance to heat stress and fungal penetration, both elevated risks during the dense humid canopy conditions of late flower.
A concentrated seaweed extract sitting alongside the standalone Scottish Seaweed powder — two forms of the same seaweed family, working at different speeds. The extract is fast-acting: cytokinins, auxins and gibberellins released within days of watering in, supporting flower initiation and reducing flower drop in the first weeks of bloom. The whole-plant powder releases slowly over the cycle. Two seaweed sources working together give a steady biostimulant signal from early flower through to finish.
Bloom 2-8-4 is a granular dry amendment. Sprinkle it on the soil surface as a top dress and water it in. No pH pens, no EC meters, no mixing reservoirs. The biology does the work. Bulk density approximately 1 g/ml — useful for converting between grams and millilitres on the kitchen scales.
Sprinkle evenly on the soil surface around the base of the plant. Water thoroughly to begin microbial breakdown. For a standard 20-litre pot, that is roughly 3–7 tablespoons per top dress. Start at 2–3 ml per litre and work up to 4–5 ml for large plants with heavy flower loads. Use our fertiliser calculator for exact quantities.
Scatter evenly and rake lightly into the top 5–10 cm of soil. Water deeply after application. Use 100–200 g for light-to-moderate feeders and established beds. Use 200–300 g for heavy feeders, fruit trees and roses during peak bloom. Outdoor beds hold nutrients longer than containers — the larger soil volume, deeper biology and natural buffering capacity mean a 4–6 week cycle gives steady nutrition without accumulation risk.
Coco coir and other soilless substrates are biologically inert — unlike soil, they contain no microbial life to break down organic matter. Add worm castings or quality compost at 20–30% of the mix to introduce the biology that makes dry amendments work. Pre-buffer with cal-mag solution before amending, or use a pre-buffered coco — unbuffered coir locks out calcium and magnesium through cation exchange. Use the upper end of the dosage range and top dress more frequently than in soil. Do not water to heavy run-off — run-off flushes dry amendments out of the root zone. Water thoroughly but stop before significant drainage.
Bloom 2-8-4 carries enough calcium and magnesium for most growers — around 8% CaO and 1% MgO from gypsum, kieserite, micronised rock phosphate and volcanic rock dust. For luxury-level results — heavy crops, container growing under high light, or growers chasing maximum yields and flower density — supplement with Dr Forest Cal-Mag at 10% of your Bloom 2-8-4 rate. Apply 10 g of Bloom, add 1 g of Cal-Mag alongside it. Match the intervals to your bloom top dress. The supplement lifts calcium and magnesium from adequate into optimal, which is where the gains in cell wall integrity, terpene production and finishing weight sit.
Use dechlorinated water. Chlorine and chloramine kill the beneficial soil microbes that break down organic matter and deliver nutrients to the plant. Stand tap water for 24 hours, run it through a carbon filter or add a dechlorinator. This single step makes the biggest difference to how well organic dry amendments perform.
Organic growing asks for a different watering approach to synthetic feeds. Water thoroughly but not to heavy run-off — wet the entire root zone, but excessive drainage flushes dissolved nutrients and microbial by-products out of the substrate. In soil, water until you see the first drops of drainage and stop. In coco coir and other soilless media this matters even more — heavy run-off washes dry amendments straight through. Let the top layer dry slightly between waterings to maintain oxygen in the root zone and encourage healthy microbial activity.
Phosphorus deficiency during flower shows as dark purple stems and petioles, slow bud development and premature lower leaf drop. Top dress at the higher end of the range (4–5 ml per litre) and tighten the interval to every 2 weeks. Some lower leaf yellowing during mid-to-late flower is normal as the plant redirects nitrogen to flowers — but if it accelerates rapidly, the plant is asking for more.
If the plant is producing new leaf growth around flower sites rather than swelling buds, nitrogen is too high. Less likely with Bloom 2-8-4's reduced 2% N, but it can happen if top dressing too frequently or at high rates. Stretch the interval to every 3–4 weeks or skip an application.
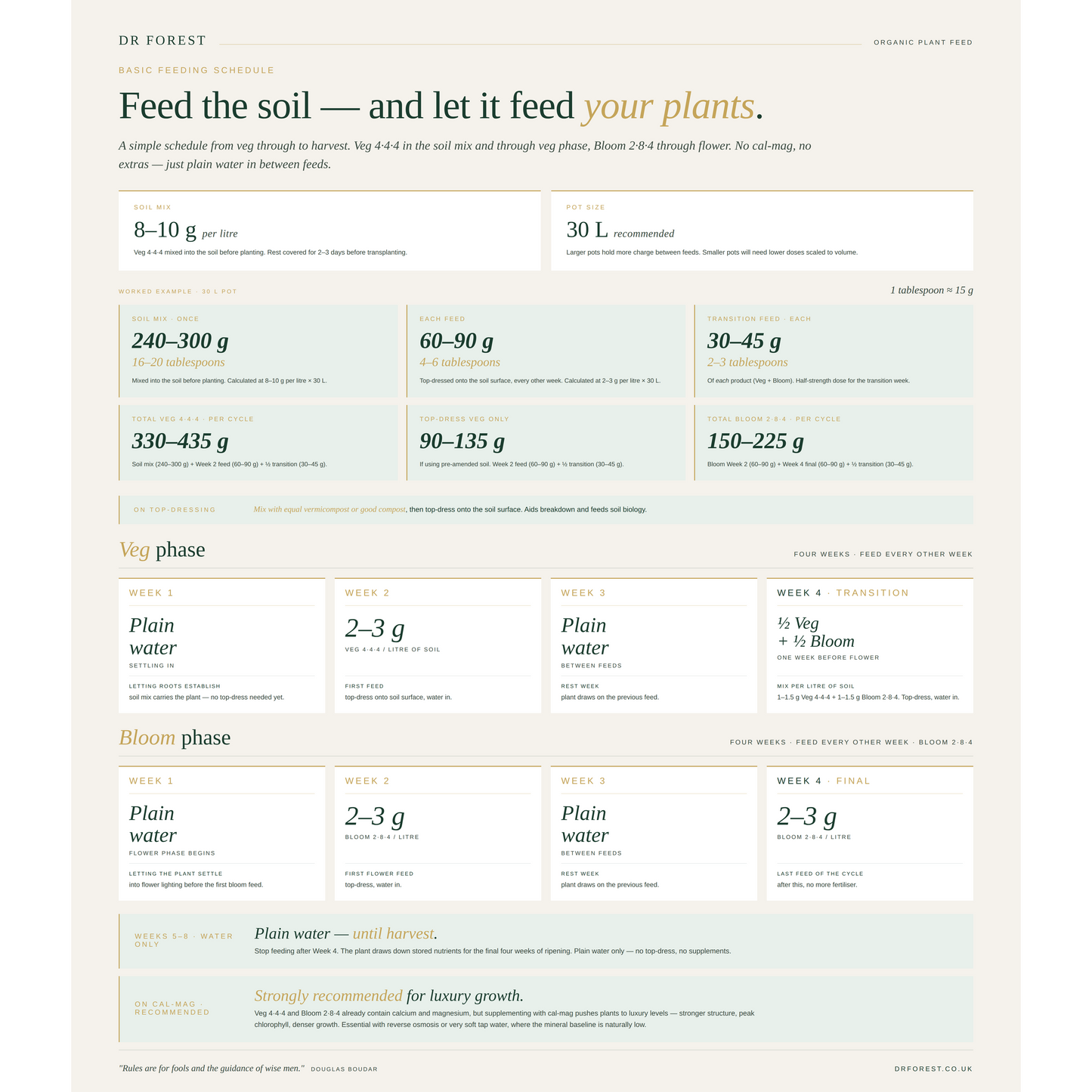
Use Dr Forest Cal-Mag through the grow to lift calcium and magnesium from adequate into optimal. Apply Amino Acid Powder as a watered-in or foliar boost during the first three weeks of flower — pre-formed nitrogen units skip the metabolic cost of nitrate conversion. Add Seaweed Powder as a fortnightly foliar to maintain biostimulant activity through flower. Top up with Dr Forest Mineral Mix for trace mineral diversity in long fruiting seasons. See our feeding schedule for the full programme, or our measurement converter to switch between grams, ml and tablespoons.
Mix in a well-ventilated area. Wear a dust mask when handling the dry powder. Wash hands after use. Keep out of reach of children and pets. Store sealed in a cool, dry place.
Bloom 2-8-4 works across every kind of flowering and fruiting plant, but the rate, frequency and timing vary depending on what you are growing. This is a working guide to which plants benefit most, what to adjust for each type, how protected and outdoor growing differ, and what to watch for through the UK season.
Tomatoes, peppers, chillies, courgettes, cucumbers, aubergines, squashes. These crops shift from leaf-building to fruit-set in a clear window — from the first flower onwards, the demand swings hard toward phosphorus, potassium and calcium. Bloom 2-8-4 hits all three. Top dress when the first flowers appear, then every 3–4 weeks through the harvest.
Strawberries, raspberries, blackberries, blueberries, apples, pears, plums, cherries. Apply once at bud-break (March–April), again at petal-fall, and a final dose at fruit-set. For strawberries, work into the soil before mulching with straw. For trees, apply to the drip line rather than the trunk. The slow-release fraction carries through ripening without spiking nitrogen during the sugar-loading phase that drives flavour.
Roses, dahlias, sweet peas, cosmos, sunflowers, peonies, fuchsias. Roses respond particularly well — apply at first bud sighting, then every 4–6 weeks through the repeat flush. For dahlias and sweet peas, start when the first buds form; the high potassium drives flower count and stem strength.
Growers who split veg and bloom across two distinct fertilisers. Bloom 2-8-4 is the primary flowering-stage feed: top dress when flowers initiate, continue through harvest, stop 2–3 weeks before cut. Pair with Veg 4-4-4 for the vegetative phase. See the transition steps in the How to Use tab.
Long fruiting seasons strip nutrients faster than the average bedding plant. These crops will not stop demanding phosphorus and potassium until you cut them down. Top dress on the higher end of the range and keep the interval tight. Pair with foliar seaweed every fortnight for biostimulant activity through the harvest.
Repeat-flowering roses take roughly six weeks from a hard prune or deadhead to the next flush. Apply at bud-break in spring, after each main flush, and a final dose in mid-August — three to four feeds across a season. Hold off from September onwards so the plant can harden off before winter. Hybrid teas and floribundas are heavy feeders; English shrub roses sit in the middle; once-flowering ramblers and species roses need only a single application at bud-break.
Hardy geraniums, asters, salvias, lavender and most Mediterranean herbs. One application at bud-break, one mid-season. Overfeeding ornamentals produces lush foliage at the expense of flower count — a common mistake. Keep the rate light.
Containers hold a finite reserve, so a small soil volume runs out of nutrients quickly under heavy bloom. Top dress every three weeks once the first flowers appear. For hanging baskets and patio pots through midsummer, this often means weekly to fortnightly when paired with a foliar seaweed.
Keep soil consistently moist but not soggy. The microbial activity that breaks down a fresh top dress needs water but also oxygen — waterlogged soil shuts down both. Water in the morning so the surface dries before evening.
Plant water demand peaks here — heavy flowers and fruit are mostly water. Allow the top inch to dry between waterings to keep oxygen in the root zone. Long deep waterings beat frequent shallow ones.
Pull back slightly. A small water deficit in the last fortnight concentrates sugars and improves flavour in fruiting crops. Do not stress the plant — wilting damages yield — but do not pour water on if it doesn't need it.
| Month | What to do |
|---|---|
| March | Hold off. Soil below 10°C means dormant microbes — no nutrient cycling yet. Top dress beds with Veg 4-4-4 if amending for spring sowing. |
| April | Bud-break for apples, pears and roses. First bloom top dress now. Strawberries beginning to flower under cloches or in protected beds — feed at flower-set. |
| May | Outdoor strawberries and soft fruit at flower-set get their feed. Greenhouse tomatoes and peppers begin bloom feeding as the first truss flowers open. |
| June | Peak feeding window. Tomatoes through second to fourth truss. Roses after the first flush — top dress straight after deadheading. Dahlias and sweet peas as buds form. |
| July | Tight intervals for protected fruiters (every 2–3 weeks). Outdoor beds settle to every 4 weeks. Watch for blossom-end rot in hot dry spells — water consistency matters more than feed. |
| August | Final rose dose by mid-month. Outdoor tomatoes still cropping. Last application for hardy perennials — peonies and asters for the late flush. |
| September | Final feeds for chillies, peppers and outdoor tomatoes still ripening on. Stop feeding ornamentals so they can harden off for winter. |
| October | Late apple and pear varieties still drawing nutrients — final dose at the drip line. Stop feeding everything else. |
| Nov–Feb | Hold off. Mulch over beds with leaf mould or garden compost. Resume bloom feeding when soil warms above 10°C — usually April. |
The blend carries ~8% calcium — soil calcium is rarely the underlying problem. Blossom-end rot is almost always an uptake issue caused by drought-flood swings or excess potassium. Water consistently. Mulch heavily. Avoid sudden flushes after a dry spell. If the issue persists, foliar calcium gives a faster response than soil applications.
Above 30°C, pollen viability drops sharply in most fruiting crops. Shade in extreme heat. The seaweed inputs and volcanic rock dust supply trace boron for pollen tube growth, but persistent drop usually points elsewhere — pollinator activity, temperature, or a sudden humidity swing. Hand-pollinate tomatoes by gently tapping the truss every other morning, or open polytunnel doors and vents more.
If switching from a high-N feed straight into early bloom and seeing sparse buds, the residual nitrogen is suppressing flower initiation. Hold off any further N supplementation and apply Bloom 2-8-4 at the higher end of the range. The reduced 2% N in the blend won't add to the problem.
Gradual yellowing from the bottom up through mid-to-late flower is the plant moving mobile nutrients (N, P, K, Mg) into flowers. This is normal and not a problem. Rapid yellowing with leaf drop in upper canopy suggests true deficiency — top dress and add a foliar seaweed.
Silica meal in the blend supports stem strength, but light limitation produces thin etiolated growth that no fertiliser can fully fix. Stake heavy crops early. For repeat grows, increase silica supplementation through veg as well as bloom.
Soft, airy flowers usually trace back to feeding too aggressively into late flower or running a higher-N product through the finishing weeks. Drop bloom feeding 2–3 weeks before harvest. For very heavy feeders, finish on Bloom 2-4-8 — the higher K ratio sharpens the final fortnight.
The transition from vegetative to reproductive growth is the most energy-intensive phase in a plant's life cycle. Flower bud differentiation, pollen formation, fruit set and seed development each require large quantities of ATP — the energy currency built around a phosphate backbone. A phosphorus-dominant ratio ensures the plant has the ATP raw material it needs precisely when demand peaks. Reduced nitrogen at 2% prevents excess vegetative growth that competes with flower development for photosynthetic resources, and potassium at 4% drives sugar transport from leaves to flowers and supports enzyme activation throughout the reproductive process.
Phosphorous Meal supplies citric-acid-soluble phosphorus at roughly 15% P₂O₅, which plant roots unlock on demand through their own root exudates. When the plant needs phosphorus, it secretes organic acids that dissolve the meal directly in the rhizosphere. Micronised rock phosphate contributes slow-release P at 31% with significant calcium — a reserve that continues feeding flowers long after faster sources are depleted. The Amino Acid Powder and the broader organic fraction add a smaller biological P contribution that microbes mineralise over weeks. Three sources, three release mechanisms.
Sulphate of potash delivers immediately available potassium with zero chloride at 50% K₂O. The base blend contributes additional K through the slower organic fraction, and volcanic rock dust adds trace potassium as part of its 60+ element mineral spectrum. Potassium during flower is critical: it activates over 60 enzymes, regulates stomatal opening and — most importantly for quality — drives the long-distance transport of sugars, organic acids and secondary metabolites from photosynthetic leaves into developing flowers and fruit. Higher potassium availability correlates directly with terpene concentration and flavour intensity.
Bloom 2-8-4 carries sulphur from four sources — sulphate of potash, kieserite, gypsum and rapeseed meal — for a total SO₃ contribution above 5%. Sulphur is a structural component of the amino acids methionine and cysteine, and a precursor for sulphur-containing volatile compounds that contribute to aroma intensity. It is also required for the synthesis of coenzyme A, which drives the mevalonic acid pathway — the primary biosynthetic route for terpenoid production in plants.
The blend carries calcium from three sources — gypsum (23% Ca), micronised rock phosphate (49% Ca) and Phosphorous Meal (9% CaO) — landing close to 8% total CaO. Magnesium comes from kieserite (~25% Mg) plus smaller contributions from the organic fraction. This is enough calcium and magnesium for the great majority of grows. For luxury-level results — heavy crops, container growing under high light, or growers pushing for maximum density and finishing weight — supplement with Dr Forest Cal-Mag to lift these structural nutrients from adequate into optimal.
The quality difference between organically and synthetically grown crops is measurable and repeatable, and it is explained by biochemistry rather than ideology. Peer-reviewed research shows that organic nutrient management produces higher secondary metabolite concentration, better flavour compound profiles and healthier soil biology — all of which compound over repeated cycles.
When soil microbes mineralise organic matter, they produce enzymes, organic acids, amino acids and volatile compounds as by-products. Many of these secondary metabolites are absorbed by plant roots and contribute directly to terpene biosynthesis, flavour intensity and aroma complexity. Synthetic feeds bypass this biology — the nutrients arrive, but the metabolic by-products that drive quality are absent.
Li et al. (2024) found that all 21 starch and sucrose metabolism genes were upregulated under organic fertilisation. Higher sugar content translates directly to improved flavour, aroma and resin production. These are the biochemical pathways responsible for the quality difference between organically and synthetically grown crops.
Cardarelli et al. (2023) reported that organic fertilisation reduces tissue nitrate concentrations by 27–50% compared to synthetic nitrogen sources. Lower tissue nitrate means cleaner, smoother material with reduced harshness — a quality parameter that matters to growers producing high-value crops for consumption.
Wang et al. (2023) synthesised 7,859 data pairs and concluded that combined organic-mineral fertilisation produces the highest crop quality outcomes of any fertiliser strategy. Bloom 2-8-4 is precisely this — organic nitrogen sources (alfalfa, frass, rapeseed, amino acids) combined with mineral nutrient carriers (gypsum, kieserite, rock phosphate, volcanic rock dust, sulphate of potash).
Ferro et al. (2022) demonstrated that organic fertiliser management increases soil organic carbon by 12.9% compared to mineral-only inputs. In living soil systems, soil carbon feeds the microbial communities that produce the metabolites driving crop quality — a compounding benefit that grows with each cycle.
Liu et al. (2021) measured urease activity +38.3%, β-glucosidase activity +122.4%, and yield increases of 15–20% under organic fertiliser management. These enzymes are the machinery of nutrient cycling — more enzyme activity means faster, more complete mineralisation of organic inputs and greater secondary metabolite production.
Shen et al. (2024) showed that unbalanced fertilisation causes 23–31% loss of actinobacterial diversity in soil. Actinobacteria produce antifungal compounds critical for disease suppression during the dense, humid conditions of late flower. Bloom 2-8-4 maintains the microbial diversity that protects the crop through its most valuable stage.
Xu et al. (2024) analysed 537 experiments and reported that organic fertilisation increased plant biomass by 56% while maintaining biodiversity. Inorganic fertilisation achieved 42% biomass increase but at the cost of soil biodiversity loss. In living soil systems, biodiversity is the mechanism of quality production.








45% of buyers here also chose Organic Veg Fertiliser 4-4-4

Balanced 4-4-4 for strong leafy growth.
from £10.99

A 0-5-10 PK feed for the final weeks of fruiting.
from £11.00

Veg and bloom together, matched to your stage.
from £20.00